
A. Qayum ( Jinnah Post-graduate Medical Centre, Karachi. )
S.M. Yusuf ( Department of Pharmacy, University of Peshawar. )
September 1983, Volume 33, Issue 9
Original Article
Abstract
The effect of adrenaline and nor-adrenaline on the spontaneously contracting and quiescent isolated proximal and central pieces of ureters obtained from female (virgin) albino rats of the Sprague Dawley strain were studicd, Adrenaline and nor-adrenaline produced a stimulant effect on the rate and amplitude of the proximal pieces of spontaneously contracting preparations. Both these drugs induced persistaltic activity in similar quiescent preparations. Adrenaline also produced a stimulant effect on the sport aneously contracting preparations of the central portions of the ureters. In view of these findings it is. suggested that as far as the responses to adrenaline and nor-adrenaline are concerned the proximal and the central portions of the isolated rat ureter preparations behave as if they possess a preponderance of alpha (excitatory) adrenergic receptors. (JPMA 33: 223, 1983).
Introduction
Ancill et al. (1972) reported that adrenaline and nor-adrenaline produced inhibitory effect on the persistaltic activity of ureters of anaesthetized rates While conducting experiments on isolated rat ureter preparations, Raz et al. (1972) observed that adrenaline inhibited the activity of these preparations. On the other hand Tindall (1972) found that in intact anaesthetized rats nor-adrenaline could produce a stimulant effect on ureters, and Hannappel and Golenhofen (1974) reported that adrenaline excited the isolated rat ureter preparations. In view of this controversy it was thought worthwhile to investigate the effects of adrenaline and nor-adrenaline on the isolated rat ureter preparations.
Material And Methods
Animals
Female (virgin) albino rats of the Sprague Dawley strain, 3-4 months old and weighing 250-300 G, were used. These animals were reared at . the animal house of Jinnah Post-graduate Medical Centre, Karachi, and were fed on Lever brother’s mouse feed.
Experimental Procedure:
i) Setting of Isolated Preparations:
The animal under experiment was stunned by a blow on the head on the 3rd day of dioestrus and the ureters were exposed and carefully dissected free to avoid stretching. After tying its upper and lower ends each ureter was cut at the points proximal to the upper and distal to the lower knot, immediately transferred to a petri dish containing Locke’s solution No. 2 and allowed to remain in a refrigerator (5-8°C) for about 24 hours (Raz et al., 1972). Each tissue was then cut in such a way as to obtain a 3-3.5 cm long piece of the proximal portion or 1.5 cm long piece of the central portion. This piece was suspended in an isolated organ bath of 10 ml capacity containing oxygenated Locke’s solution No. 2 at 38°C, by fixing its lower end to the oxygen tube and the upper end to a force displacement transducer, and was subjected to a tension of 0.5 G. After the appearance of spontaneous activity which took about 5-30 minutes sufficient time was allowed tifi the activity became reasonably stable. Tissues which showed irregularities in their activity were not used for experiments. Some quiescent tissues (i.e. those in which spontaneous activity did not appear even after a rest for one hour), were also used for experiments; all these quiescent tissues were obtained from the proximal portion and 30-3.5 cm long pieces were suspended in isolated organ baths as done in the case of spontaneous preparations.
ii) RecordingDevice:
A Grass polygraph (Model 7B) was used for recording the activity of the tissues. After the usual adjustments (i.e. adjusting the sensitivity at 2 cm deflection of the pen when the amplifier was on “CAL” and then when the amplifier was on “USE” and preamplifier on “CAL”, the upper end of the tissue was connected through a fIne cotton thread to the force displacement transducer (Model FTO3C). The preamplifier was then adjusted on sensitivity of 0.2 MV/cm and the required amount of tension was placed on the tissue. The final sensitivity (MV/cm) was kept at 0.01.
iii) Recording of Contractile Responses:
Adrenaline and nor-adrenaline were used in concentrations of I and 10 ug/ml. Each concentration was allowed to remain in the bath for two minutes. The rate of spontaneous contractions is expressed as the number of contractions per two minutes. The amplitude of contractions has been measured in millimeters. The contraction having a contractile response with maximum amplitude was selected for measurement (Qayum and Yusuf, 1977).
Nutrient Solutions and Drugs
Locke’s solution No. 2 of the following constitution (in G/litre of distilled water) was used:- NaCl, 9.0; KC1, 0.42; CaCl2, 0.24; NaHCO3, 0.3; Dextrose, 1.0. The following drugs were used: Adrenaline Hcl (Pak Davis, U.S.A.) and Nor-adrenaline Bitartrate (Winthrop, U.S.A.); the concentrations of these drugs are expressed in terms of active base.
Results
Spontaneously Contracting Preparations
1) Proximal Portion
When used in concentrations of 1 and 10 ug/ml adrenaline and nor-adrenaline produced an immediate stimulant effect on these preparations, producing increase in the rate as well as the amplitude of spontaneous contractions. Typical responses to 10 ug/ml concentrations of adrenaline and nor-adrenaline are shown in Fig- I.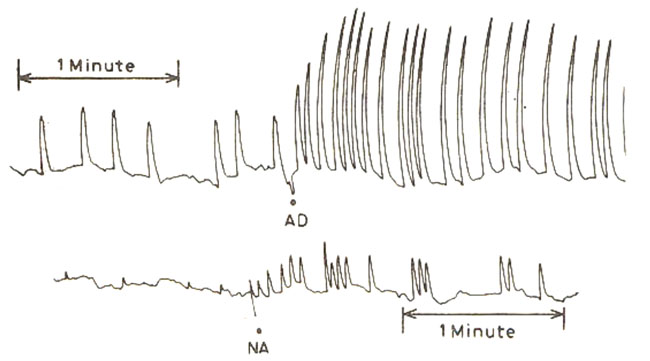
The details are as follows:-
Fig.I Effect of 10 ug/mi adrenaline (AD) and noraadrenaline (NA) on the spontaneously contracting proximal portion of the isolated rat ureter preparations.
Adrenaline (1 ug/ml):
The increase in the rate and amplitude of spontaneous contractions of 15 preparations is shown in Table-I.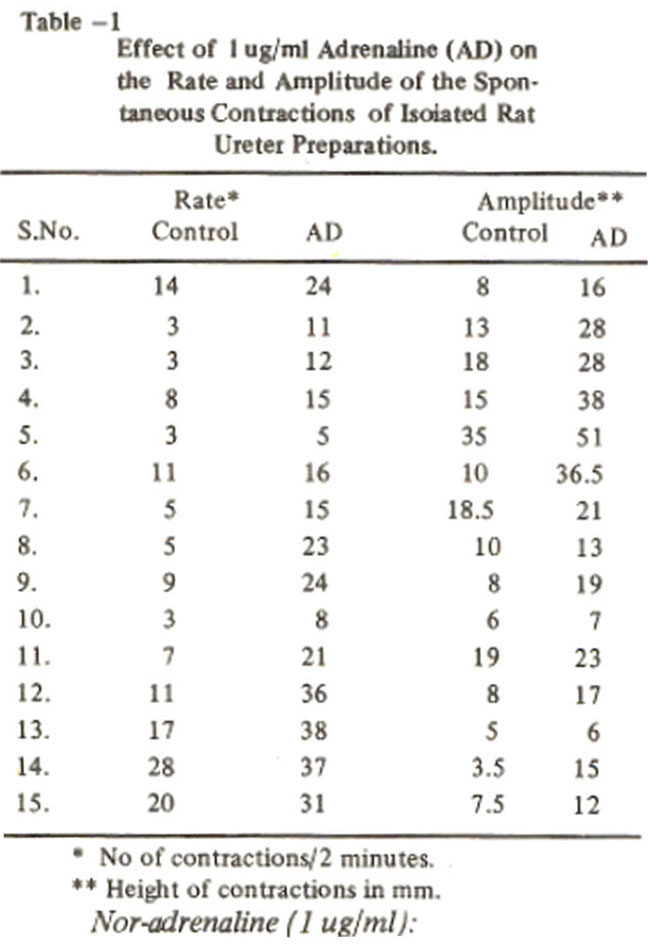
With this concentration the rate of spontaneous contractions increased from 9.8 ± 1.9 to 21.1 ± 2.8 per two minutes and the amplitude of contractile responses increased from 123 ± 2.0 to 22.0 ± 3.2 mm (Table III)
Adrenaline (10 ug/mi):
The increase in the rate and amplitude of spontaneous contractions of 13 preparations is shown in Table II.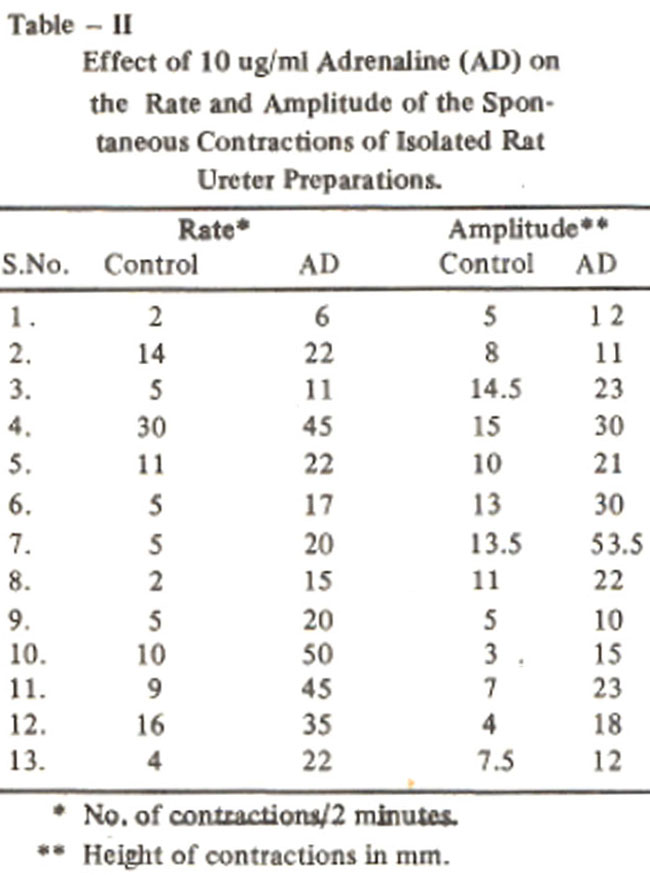
With this concentration the rate increased from 9.1 ± 2.2 to 25.4 ± 3.2 per two minutes and the amplitude of contractile responses increased from 9.7 ± 1.6 to 21.6 ± 3.2 mm (Table III).
The increase in the rate and amplitude of spontaneous contractions of 11 preparations is shown in Table IV. With this concentration the rate increased from 8.7 ± 1.4 to 18.5 ± 1.8 per two minutes and the amplitude increased from 8.4 ± 1.1 to 14.7 ± 1.6mm (Table VI).
Nor-adrenaline (10 ug/ml):
The increase in the rate and amplitude of spontaneous contractions of 8 preparations is shown in Table V.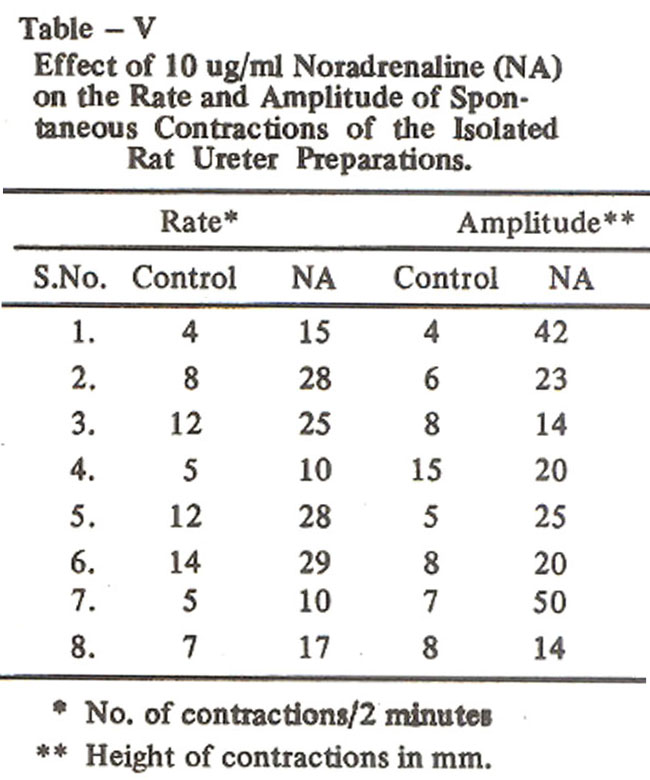
With this concentration the rate increased from 8.4 ± 1.3 to 20.3 ± 2.9 per two minutes and the amplitude of contractile responses increased from 10.5 ± 2.4 to 21.6 ± 2.8 mm (Table VI).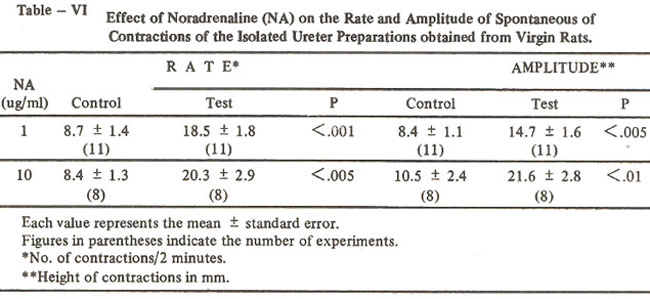
experiments with adrenaline and nor-adrena. line the values for the rate and amplitude of the induced contractions are shown in Table VII.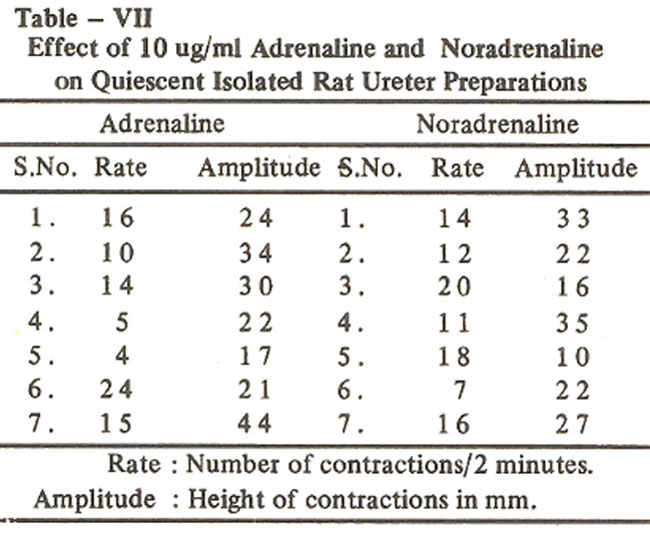
Discussion
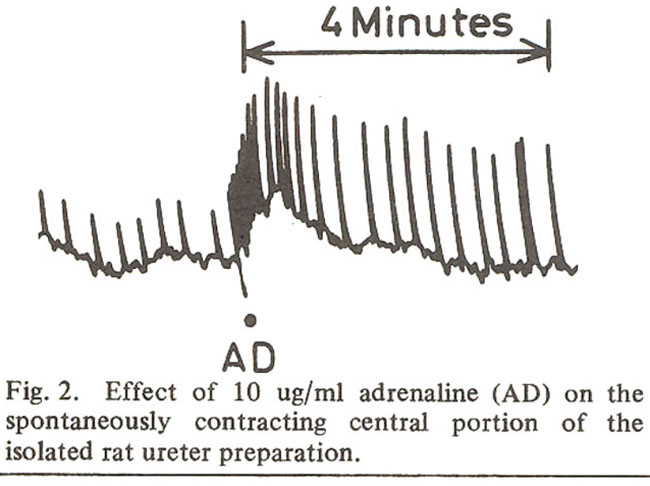
The present study revealed that adrenaline and noradrenaline (1 and 10 ug/ml), when allowed to remain in the bath for 2 minutes, produced an increase in the rate and amplitude of the proximal 3-3.5 cm long pieces of spontaneously contracting isolated preparations of ureters obtained from virgin rats in dioestrus. Both these drugs (10 ug/mI) induced spontaneous activity in similar quiescent preparations. Adrenaline also produced a stimulant effect on the spontaneously contracting preparations obtained from the central portion of the ureter.
Ancill et al. (1971) reported that the electrical stimulation of sympathetic nerves to ureters in anaesthetized rats inhibited the rate of spontaneous contractions of the ureters. These fmdings indicate that the endogenous noradrenaline, released at the sympathetic nerve endings, inhibits the activity of the rat ureter. To further evaluate these observations, Ancil et al. (1972) studied the effect of intravenously administered noradrenaline and adrenaline on the spontaneous contractions of the ureters of the anaesthetized male rats of Sprague Dawley strain. These studies revealed that both these drugs produced a decrease in the rate of spontaneous contractions of the ureters. These observations indicate that in the intact rats adrenaline and noradrenaline both have an inhibitory effect on the peristaltic activity of the ureters. On the other hand, Tindall (1972) observed that noradrenaline produced a stimulant effect on the normal peristaltic activity (rate of spontaneous contractions) of the anaesthetized rats. He found that in some experiments the effect of noradrenaline was Inhibitory, but he is of the opinion that in these preparations, unlike the normally contracting ureter, the peristaltic activity was ‘retrograde’ (i.e., instead of being initiated near the renal end, the activity appeared in a portion near the urinary bladder and then started travelling upwards), and, thereby, not normal. On the basis of these observations Tindall (1972) believes that noradrenaline produces a stimulant effect on the normal ureteric contractions through the activation of alpha excitatory adrenergic receptors. Though the observations in the present study that noradrenaline stimulated the rate as well as the amplitude of spontaneous contractions of the isolated rat ureter preparations are not in accordance with the fmdings of Ancill et al. (1972) they can be explained on the basis of the observations of tindall (1972).
Raz et al. (1972) studied the effect of adrenaline on the rate of spontaneously contracting central portions of the isolated rat ureter preparations. They found that in concentrations of 10 ug/mi adrenaline produced a decrease in the rate of spontaneous contractions of this preparation. These fmdings are in accordance with the observations of Ancill et al. (1972) who reported similar effects in the intact anaesthetized rats. Our findings that in concentrations of 1 and 10 ug/mi adrenaline produces a stimulant effect on the rate and amplitude of spontaneous contractions are, therefore, contradictory to the observations of both of these groups of workers.
The differences between the observations of Ancill et al. (1972) and those obtained in the present work can be explained on the basis of the fact that the experimental conditions in both these studies were not the same i.e., Ancill et al. (1972) studied the effect of adrenaline on the intact ureters of the anaesthetized rats, whereas, we have used isolated preparations. As Raz et al. (1972) performed experiments on 1.5 to 2 cm. long pieces obtained from the central portion of the ureter, it appeared that the above mentioned variations in the responses could also be due to variations in the experimental conditions. In view of this situation, some experiments were performed on 1.5 cm long pieces obtained from central portions of the ureters. However, in these experiments also adrenaline (10 ug/mI) produced an increase in the rate and amplitude of spontaneous contractions. Raz et al. (1972) have used female albino rats of the Lewis strain, but in the present study experiments have been performed on rats belonging to the Sprague Dawley strain. There is, therefore, a possibifity that the discrepancy in the results obtained by Raz et al.(1972) and that reported in the present work may most probably be due to variation in the strains of the rats.
It has been assumed that the rat ureter possesses both alpha excitatory and beta inhibitory adrenergic receptors (Tindall, 1972) and that adrenaline, noradrenaline and isoprenaline produce their effects mainly through the activation of these receptors (Raz et al., 1972; Tindall, 1972). On the basis of this assumption the inhibitory effects of adrenaline and noradrenaline have been attributed to the relative predominance of beta receptors In this preparation (Ancil et al., 1972; Raz et aL, 1972). However, on the basis of the results obtained in the present work it is suggested that the isolated rat ureter preparations used in this study have a preponderance of alpha (excitatory) adrenergic receptors. These conclusions are in conformity with the findings of Tindall (1972) who, by using a close arterial injection technique in the anaesthetized rats showed that the proxunal end of the ureter behaves as if it possessed alpha excitatory adrenergic receptors, and the lower end behaves as if it contained beta inhibitory adrenergic receptors. The findings in the present work that adrenaline and noradrenaline could initiate spontaneous activity even in quiescent preparations of ureters, further substantiate the observations made on the effects of these drugs on the spontaneously contracting preparations. The stimulant effect of adrenaline on the isolated quiescent preparations of the rat ureters has also been reported by Hannappel and Golenhofen (1974).
It has been observed that. adrenaline pro duces an increase in the rate as well as amplitude of spontaneous contractions of the isolated rat tenal pelvis muscle (Finberg and Peart, 1970) and pyeloureter (Hannappel and Golenhofen, 1974) preparations. The results obtained in the present study, therefore, indicate that as far as responses to adrenaline are concerned, the isolated rat ureter behaves like these preparations.
Variations in responses to adrenaline have also been shown in the case of dog ureter. Thus, Butcher et al. (1957) observed that adrenaline pro duced no effect on the rate of spontaneous contractions of the dog ureter, Mazella and Schroeder (cited by McLeod et al., 1973) reported that it produced inhibitory effect on this preparation, and Chen et al. (1957), Kaplan et al. (1968), MpLeod et al. (1973) and Hannappel and Golenhofen (1974) found it to produce a stimulant effect on the rate of spontaneous contractions of this preparation. Similarly, although the findings of several workers (Malin et al., 1968; Kaplan, 1968; McLeod et al., 1973) indicate that the dog ureter contains both alpha and beta adrenergic receptors, the experiments of other workers (Struthers, 1973) reach to the conclusion that this tissue contains only alpha adrenergic receptors.
Acknowledgements
The authors are grateful to Dr. Gut Rabman, Director, IRNUM, Peshawar, for providing typing facffities.
References
1. Ancill, R.J., Jackson, D.M. and Redfern, P.H. (1971) The effect of sympathomirnetic drugs and sympathetic nerve stimulation on the activity of the rat ureter in vivo. J. PhysioL Lond., 213:67 p.
2. Ancill, RJ., Jackson, D.M. and Redfern, P.H. (1972) The pharmacology of the rat ureter in vivo. Br. J. Pharmac., 44 :628.
3. Butcher, H.R., Sleator, W. and Schinandt, W. (1957) A study of the peristaltic conduction mechanism in the canine ureter. J.UroL, 78:221.
4. Chen, PS., Eminel, V.M., Benjamin, J.A. and Distefano, V. (1957) Studies on the isolated dog ureter; The pharmacological action of histamine, levarterenol and antihistaminics. Arch. Int. Pharmacodyn., 110:131.
5. Finberg, J.P.M. and Peart, W.S. (1970) Function of smooth muscle of the rat renal pelvis; response of the isolated pelvis muscle to angiotensin and some other substances. Br. J. Pharmac, 39:373.
6. Hannappel, J. and Golenhofen, K. (1974) Comparative studies on normal ureteral peristalsis in dogs, guinea-pigs and rats. Pflugers Arch., 348 :65.
7. Kaplan, N., Elkin, M. and Sharkey, J. (1968) Ureteral peristalsis and the autonomic nervous system. Invest. Urol., 5:468.
8. Malin, J.M. Jr., Boyarsky, S., Labay, P. and Gerber, C. (1968) In vitro isometric studies of the ureteral smooth muscle. J. Urol., 99:396.
9. McLeod, D.G., Reynolds, D.G. and Swan, K.G. (1973) Adrenergic mechanisms in the canine ureter. Am. J. PhysioL, 224:1054.
10. Qayum, A. and Yusuf, S.M. (1977) The effect of azapetine phosphate on the isolated rat ureter pre paration. Life Sci., 21:1617.
11. Raz, S., Zeigler, M. and Caine, M. (1972) Hormonal Influence on the adrenergic receptors of the ureter. Br. J. UroL, 44 :405.
12. Struthers, N.W. (1973) Experimental model for evaluating drug effects on the ureter. Br. J. UroL, 45:23.
13. Tindall, A.R. (1972) PrelimInary observations on the mechanical and electrical activity of the rat ureter. J.PhysioL Lond., 223:633.
Journal of the Pakistan Medical Association has agreed to receive and publish manuscripts in accordance with the principles of the following committees: